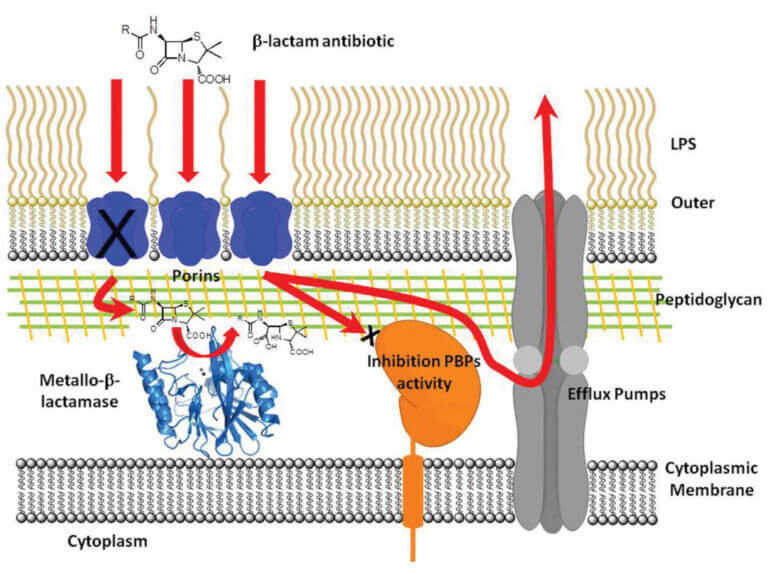
Los antibióticos β-lactámicos son los agentes quimioterapéuticos más empleados para el tratamiento de infecciones bacterianas. El principal mecanismo de resistencia a estos fármacos es la síntesis de β-lactamasas, enzimas que hidrolizan e inhiben la acción de estos antibióticos. Las metalo-β-lactamasas (MβLs) constituyen la generación más reciente de estas enzimas. Las MβLs poseen un amplio espectro de acción, ya que inactivan todos los antibióticos β-lactámicos, incluidos los carbapenemes (el último recurso empleado en la clínica).
La diseminación de genes que codifican estas enzimas en organismos oportunistas y patógenos es hoy de escala mundial. Este cuadro se agrava por la diversidad estructural de las distintas MβLs, que dificulta el diseño de un inhibidor eficiente para estas enzimas. La enzima NDM-1 (http://www.bbc.co.uk/news/health-10925411), en particular, está siendo diseminada en todo el mundo rápidamente, y no se cuenta con inhibidores de uso clínico para estas enzimas, lo que representa una amenaza para la salud mundial.
Nuestro objetivo es elucidar la relación estructura-función de estas enzimas mediante estudios bioquímicos, estructurales, mecanísticos y evolutivos, con el fin último de diseñar un inhibidor de aplicación clínica. Hasta el momento, hemos logrado: (1) proponer un mecanismo catalítico común para las MβLs a partir de la caracterización de un intermediario de reacción, (2) identificar la especie funcional in vivo de las mismas y (3) explorar sus posibles mecanismos evolutivos. En la actualidad, utilizamos estos conocimientos previos para diseñar y generar inhibidores de MβLs, bajo la hipótesis de que, a pesar de su diversidad estructural, las MβLs actúan utilizando el mismo mecanismo catalítico.
Nuestro grupo lleva adelante un estudio interdisciplinario, usando técnicas de biología molecular, bioquímica, biología estructural y enzimología moderna. Los estudios mecanísticos se realizan usando técnicas de mezcla rápida y varias espectroscopías para seguir cambios en el sitio activo durante el recambio catalítico en la escala de tiempo de los milisegundos, con el propósito de identificar intermediarios de reacción.
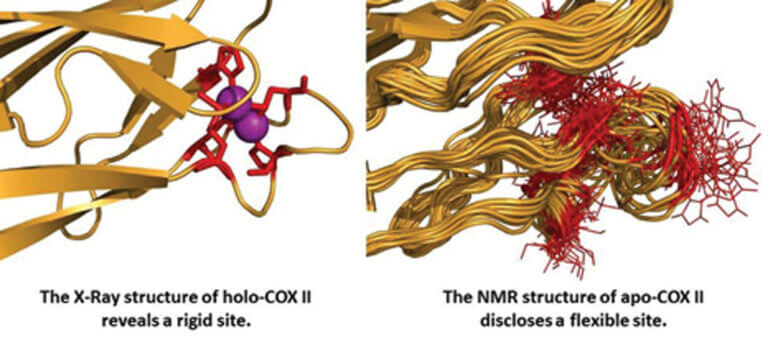
Utilizamos cristalografía y/o Resonancia Magnética Nuclear (RMN) para caracterizar el modo de unión de posibles inhibidores a estas enzimas. También usamos estrategias de evolución dirigida in vitro como un medio para pronosticar la evolución de estas enzimas en ambientes hospitalarios, y para entender de qué modo se fijan las mutaciones en el proceso evolutivo.
Director de Grupo: Vila, Alejandro